
In today’s society, an overconsumption of fats, especially the unsaturated trans fats, significantly increases the risk of developing lifestyle diseases such as obesity and dyslipidemia.
Meanwhile, interest in the anti-obesity and prebiotic properties of seaweed polysaccharides has been steadily growing. The potential of Saccharina japonica polysaccharides as anti-obesity and prebiotic agents is significant, though the underlying mechanisms remain unclear.
In this blog, I would like to share the following study, “Metagenomic Insights into the Anti-Obesity Effect of a Polysaccharide from Saccharina japonica” by Ying Song et al. The study examined the potential of S. japonica polysaccharide fraction (SjC) to prevent high-fat diet-induced obesity in mice, focusing on its underlying mechanism.
First, Saccharina japonica polysaccharides (SjC) were characterized as a sulfated polysaccharide of 756 kDa and randomly divided into three groups (control, model, and SjC groups). Mice in the model and SjC groups were fed a high-fat diet (HFD diet), while mice in the control group were fed a low-fat diet. The experimental design involved providing the SjC group with drinking water supplemented by a 1 mg/mL SjC solution, freely available for 16 weeks; in contrast, the control groups had ad libitum access to regular drinking water throughout the same timeframe.
The study assessed the effects of administering polysaccharide SjC for 16 weeks to C57BL/6J mice with high-fat-diet-induced obesity and metabolic disorders. The results showed that 16 weeks of HFD intake significantly increased the body weight of mice compared to the normal group, but SjC supplementation effectively prevented HFD-induced weight gain. SjC also effectively reduced the energy efficiency of mice. The experiment showed that the high-fat diet (HFD) caused a parallel increase in the epididymal fat and liver weight in the mice; this increase, however, was successfully alleviated by treatment with SjC. High-fat diets (HFD) caused significantly increased glucose and insulin resistance in mice versus controls, an effect partially reversed by SjC supplementation, which significantly improved insulin resistance and slightly improved glucose tolerance. Additionally, SjC slightly alleviated the changes in serum total cholesterol and triglyceride levels caused by HFD, but the difference was not statistically significant.
The effect of SjC supplementation on the gut microbiota composition of HFD-fed mice was investigated by 16S rRNA gene sequencing of cecal contents. SjC administration slightly decreased the α diversity index (Chao 1) of gut microbiota compared with the model group, but the difference was not significant. SjC administration mainly increased the relative abundance of Desulfitobacteria and decreased the relative abundance of Proteobacteria at the phylum level. PCA plots of taxa at the species level showed that the gut microbiota composition of mice in the HFD group was significantly different from the other two groups, and SjC supplementation changed the gut microbiota structure of HFD mice to that of normal diet-fed mice, especially on the PC1 axis. A comprehensive analysis of gut microbiota composition, utilizing 16S rRNA gene sequencing to construct a phylogenetic tree, identified 214 bacterial species in total; notably, the relative abundance of 38 of these species displayed statistically significant differences across the three groups analyzed.
It is important to note that a subset of the 214 bacterial species, specifically 14, displayed high abundance and a significant positive correlation with the addition of polysaccharides to the system. HFD feeding increased the relative abundance of Desulfuro fairfieldensis, Clostridium sp., Dorea sp., Christensenella minuta, Turicibacter, and Akkermansia compared with the normal group, while SjC supplementation significantly suppressed the HFD-induced increase of these six species. Furthermore, SjC intervention significantly suppressed the relative abundance of Coriobacteriaceae UCG-002 and increased Muribaculaceae compared to the normal group.
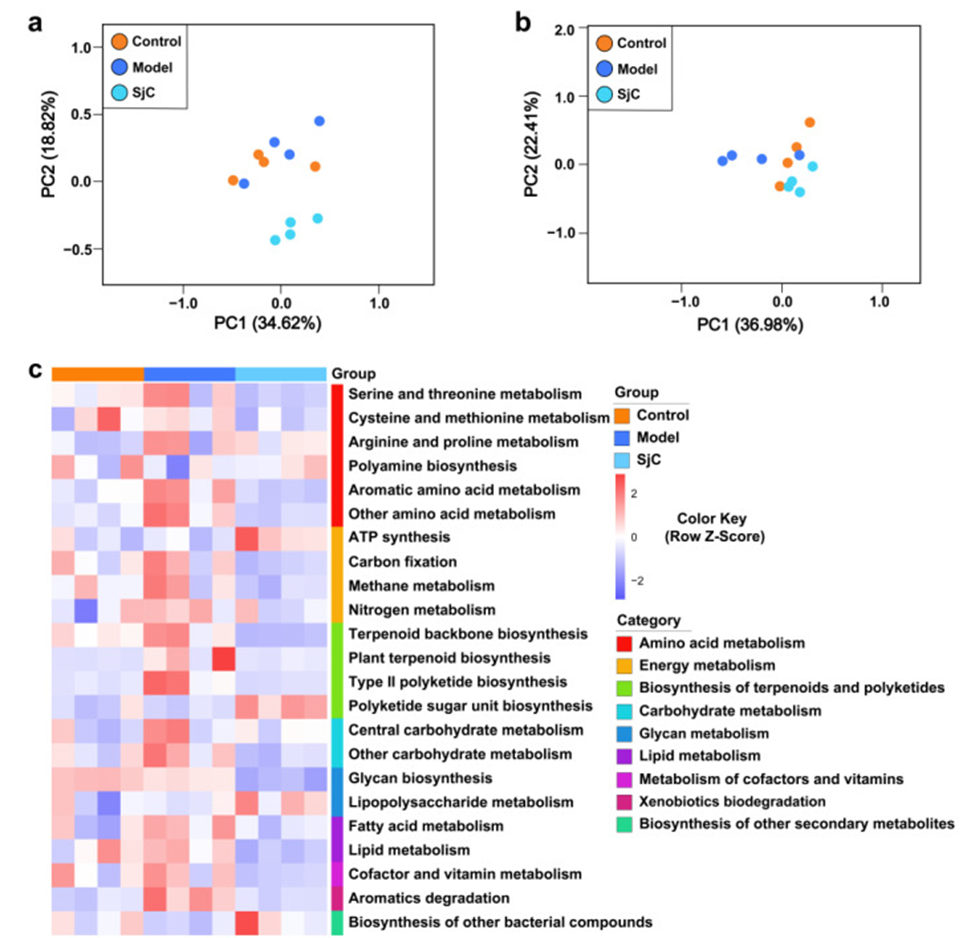
The influence of polysaccharide SjC on gut microbiota was investigated through shotgun metagenomic sequencing, focusing on functional gene and metabolic pathway abundance. As shown in Figure 1a, the unsupervised PCA plot showed that the composition of gut microbiota functional genes in the control group was highly overlapped with that in the model group, whereas the SjC group was largely separated from the other groups, suggesting that SjC supplementation significantly altered the composition of gut microbiota functional genes. Figure 1b shows a PCA plot of metabolic pathways; the SjC group clustered with the control group, but the model group was clearly distinct from both along the PC2 axis.
SjC supplementation, while affecting the abundance of certain functional genes, did not disrupt the metabolic homeostasis of the gut microbiota. Metabolic pathway enrichment analysis revealed pathways that were significantly altered among these three groups (Figure 1c). In mice fed an HFD, most amino acid metabolic pathways were enhanced, and SjC supplementation suppressed the metabolic levels of these amino acids in the gut microbiota. This trend was also seen in some metabolic pathways in energy metabolism and lipid metabolism. The addition of SjC enhanced metabolic pathways like ATP synthesis, the biosynthesis of polyketide sugar units, and the metabolism of lipopolysaccharides. SJC supplementation also decreased glycine biosynthesis.
The attempt successfully resulted in connecting the six MAGs with 16S rRNA gene sequences. The draft genome of Akkermansia suggested that it harbors 2475 protein-coding genes, of which 7.1% were annotated as carbohydrate-active enzymes. The study also revealed that seven additional gut bacterial species prevalent in SjC contain a significant number of genetic loci for polysaccharide use. A significant correlation was observed between mouse body weight and the relative abundance of six tentatively identified bacterial strains—including Desulfurovibrio (bin 33), Akkermansia (bin 80), Muribaculum (bin 271), and an uncultured member of the Muribaculaceae family (bins 120)—suggesting a potential combined role in the development of obesity.
The study concluded that SjC, a polysaccharide fraction from ‘S. japonica’, successfully countered obesity, metabolic problems, and gut dysbiosis induced by a high-fat diet. The main prebiotic action of SjC is regulating the proportions of Desulfurovibrio and Akkermansia bacteria and suppressing amino acid processing within the gut microbiome. SjC’s anti-obesity effects might stem from changes in the gut bacteria responsible for fucoidan degradation, according to the findings.
Source: Foods. 2023 Feb 3;12(3):665. doi: 10.3390/foods12030665