The skin is the largest organ in the human body and acts as a barrier to protect our body from external particles, chemical substances, and ultraviolet (UV) irradiation. UV irradiation by sunlight is believed to be a major environmental factor that causes skin damage through a photoaging reaction, which also produces an enzyme called “gelatinase. Gelatinase is also called “wrinkle egg” when exposed to the harmful ultraviolet rays. It causes damage to the epidermis and denatures collagen in the dermis, increasing the incidence of wrinkles in the future.
Thus, it causes damage to the epidermis and denatures collagen in the dermis, increasing the incidence of wrinkles in the future. It can also develop into skin cancer. Melanin, a skin pigment, helps prevent UV-induced skin damage, but overproduction and accumulation can cause pathological or cosmetic problems. On a positive note, fucoidan, a polysaccharide in brown algae, has been reported to be helpful as a cosmetic ingredient for its hyaluronic acid-degrading enzyme action inhibitory activity and antioxidant action. Therefore, in this blog, I would like to share the effect through the research of “Anti-photoaging and anti-melanogenesis effects of fucoidan isolated for Hijiki fusiforme and its underlying mechanism” by Lei Wang et al.

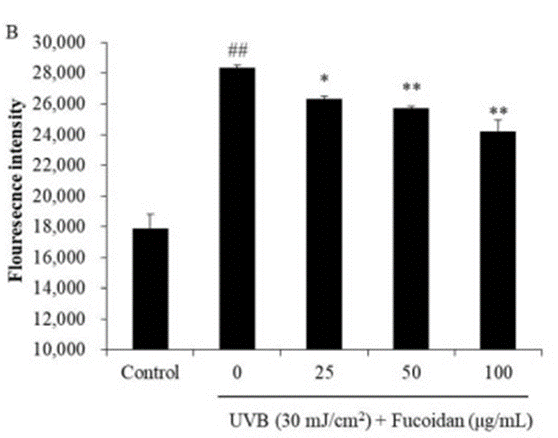
Initially, they verified the anti-photoaging effect and melanin production inhibitory effect of fucoidan extracted from Hijiki. The anti-photoaging result of fucoidan was verified using the human epidermal keratinocyte cell line HaCaT. The mechanism by which photoaging occurs is the overproduction of reactive oxygen species (ROS). UV-B-irradiated HaCaT cells significantly induced ROS production compared to untreated controls, while the addition of fucoidan reduced ROS production in a dose-dependent manner. To investigate the effects of fucoidan in more detail, they measured the formation of apoptotic bodies and the expression of apoptosis-related proteins in UV-B-irradiated HaCaT cells.
As a result, the rate of apoptotic body formation decreased in a dose-dependent manner in fucoidan-supplemented cells (Fig. 1). In addition, in HaCaT cells irradiated with UV-B, the protein that induces apoptosis of skin cells (Baz, Caspase3) increased significantly.
On the contrary, the anti-apoptotic protein'(Bcl-xL, PARP) decreased significantly. Thus, by adding fucoidan, those abnormal increases in Baz, Caspase-3, and decreases in Bxl-xL, PARP were alleviated to near normal. This report suggests that fucoidan regulates apoptosis-related signals by suppressing intracellular ROS production associated with UV irradiation and protects HaCaT cells from skin photoaging. Next, fucoidan’s melanin production inhibitory effect was examined using the malignant melanoma cell line B16F10. An enzyme called tyrosine kinase (Tyr) is involved in the melanin reaction.
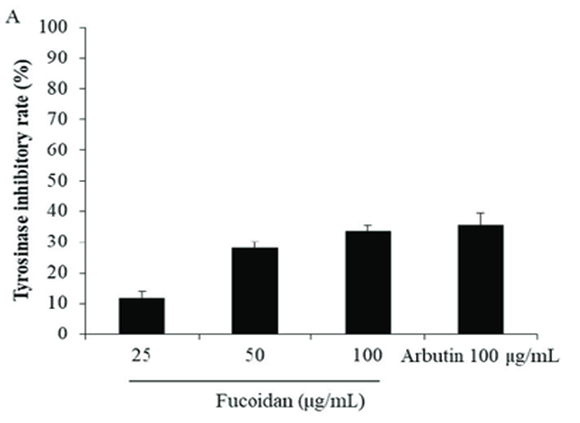
Hence, when the Tyr activity inhibitory effect of fucoidan was evaluated, the Tyr activity inhibitory rate by fucoidan was 33.6% at a high concentration (100 µg / mL). It indicates that it has a physiological effect comparable to arbutin, known for its whitening effect. (Fig. 2).
In addition, the activity of Tyr-related proteins (TRP-1, TRT-2) involved in melanin biosynthesis and ERK engaged in the expression and induction of MITF was verified using B16F10 cells treated with the melanocyte-stimulating hormone α-MSH. It turns out that fucoidan works on controlling it. These studies suggest that fucoidan has a skincare effect that suppresses photoaging and melanin production in skin cells and is expected to be applied to fucoidan as a raw material for pharmaceuticals and medicated cosmetics.